 Øyets utvikling
Øyets utvikling
Oversatt herfra.
Øyets utvikling har alltid vært et dilemma for evolusjonister fra Darwins tid til i dag. Selv om Darwin, Richard Dawkins og andre evolusjonister har forsøkt å forklare hvordan et øye kunne utvikle seg, er deres løsninger klart utilfredsstillende. Det finnes mange slags øyne, men det kan ikke produseres noen øyeformasjoner fra enkle til komplekse i den naturlige eller fossile verden. Videre er det enkleste øyet, øyestedet, ikke et øye, men pigmenterte celler som brukes til fototakser (mot/til -lyset); likevel krever det en enormt kompleks mekanisme for å fungere som et visjonssystem.
Bilde 1. Darwin realistisk
Det sammensatte øye til et insekt
 Figur 1. Det sammensatte øye av et insekt. Merk at øyet består av hundrevis eller flere separate øyne som på noen måter er mer komplekse enn det menneskelige øye. (Etter Mitchell et al .). 48
Figur 1. Det sammensatte øye av et insekt. Merk at øyet består av hundrevis eller flere separate øyne som på noen måter er mer komplekse enn det menneskelige øye. (Etter Mitchell et al .). 48
Konseptet med Ikke-Reduserbar Kompleksitet (IRK) er blitt et viktig verktøy i intelligent design teori. Et av de beste eksemplene på IK er utformingen av dyreøyet. Øyene er kritiske fordi, for "de aller fleste dyr", er visjon deres "viktigste link til verden". (1) Darwin anerkjente levende problemet med øyeevolusjon og den alvorlige hindringen som det var for hans teori. I hans ord:
"For å anta at øyet, med all sin unike mekanismer for å justere fokuset til forskjellige avstander, kunne tillate forskjellige mengder lys og for korreksjon av sfærisk og kromatisk avvik, kunne ha blitt dannet ved naturlig utvalg er, innrømmer jeg fritt, absurd i høyest mulig grad. ' (2)
Ikke desto mindre følte Darwin at det tilsynelatende uoverstigelige problemet med utviklingen av det han kalte et organ av ekstrem perfeksjon og kompleksitet, kunne løses. (2) Han inkluderte et tre-siders forslag om mellomliggende stadier der øynene kunne ha utviklet seg gjennom gradvise skritt. (3) Disse trinnene inneholdt følgende:
1. lysfølsom celle 2. aggregater av pigmentceller uten nerve 3.en optisk nerve omgitt av pigmentceller og dekket av gjennomsiktig hud 4. pigmentceller danner en liten depresjon og deretter en dypere depresjon 5. huden over depresjonen tar gradvis en linseform 6. utviklingen av muskler som lar linsen justere seg.
Disse stadiene i levende dyr antas å utgjøre store bevis for øyets utvikling. (4) Isaak hevder at alle disse trinnene er mulige fordi alle finnes i dyr som lever i dag:
'Inkrementene mellom disse trinnene er små og kan brytes ned i enda mindre trinn. Naturlig utvalg bør under mange omstendigheter favorisere trinnene. Siden øynene ikke fossiliserer godt, vet vi ikke at øyets utvikling fulgte akkurat den veien, men vi kan absolutt ikke hevde at det ikke finnes noen vei. (5)
University of Chicago biologi Professor Jerry Coyne skrev at menneskelige "... øyne opptrådte ikke plutselig som fullverdige kameraøyne, men utviklet seg fra enklere øyne, med færre komponenter, i forfedre. Darwin adresserte briljant dette argumentet ved å kartlegge eksisterende arter for å se om man kunne finne funksjonelle, men mindre komplekse øyne som ikke bare var nyttige, men også kunne bli knyttet sammen i en hypotetisk rekkefølge som viser hvordan et kameraøye kan utvikle seg. Hvis dette kunne gjøres - og det kan - forsvinner argumentet for ikke-reduserbar kompleksitet, for øynene til eksisterende arter er åpenbart nyttige, og hvert trinn i den hypotetiske sekvensen kan således utvikles ved naturlig utvalg. (6)
Det eldste øyet i fossilregistreringen, den av en trilobitt, er et meget komplekst fasettert sammensatt øye som "daterer" tilbake til Kambrium, som vanligvis dateres for ca 540 millioner år siden.
Den dominerende teorien ble skissert av Dennett, som konkluderte med at alt øyeevolusjon krever er '... sjeldne ulykker som gir et heldig dyr en mutasjon som forbedrer visjonen over sine søskens; Hvis denne forbedringen hjelper det å få flere avkom enn sine rivaler, gir dette evolusjonen en mulighet til å heve lista og et reisverk (ratchet) for øyets design med ett tankeløst skritt. Og siden disse heldige forbedringene akkumuleres - dette var Darwins innsikt-kan øyne automatisk bli bedre og bedre og bedre uten noen intelligent designer. ' (7)
Andre er ikke så selvsikre. Melnick konkluderte med at øyet er et vidunder, og at "dens enorme kompleksitet og mangfold i naturen, så vel som sin skjønnhet og perfeksjon i så mange forskjellige skapninger i verden, trosser forklaring selv av makroevolusjonens flinkeste supportere." (8) Denne artikkelen utforsker disse motstridende synene.
Bilde 2. Øyet hos virveldyr og bløtdyr
 Evolusjon av øyet
Evolusjon av øyet
Avansert visjon vises nesten helt i begynnelsen av fossilregistreringen. Det eldste øyet i fossilregistreringen, det av en trilobitt, er et meget komplekst fasettert sammensatt øye som "daterer" tilbake til Kambrium, som vanligvis dateres for ca 540 millioner år siden. (9,10) Det fossile beviset viser at fra begynnelsen av fossilregistreringen er øynene svært komplekse, høyt utviklede strukturer. Vi har også 'levende fossiler', dyr som har vært nesten uendret siden tidlig i historien. Biolog ved universitetet i Salford, Laurence R. Croft, skrev at den "nøyaktige opprinnelsen til virveldyrøyne er fortsatt et mysterium. Den fascinerende tingen om øyets utvikling er dens tilsynelatende plutselige oppdukking. ' (11) Spesielt viser fossilene at visjonen oppsto "i den tidlig kambriske-tid", som darwinister setter til ca. "for 530 millioner år siden". (12)
Videre, selv om de "kambriske dyrene ikke var den samme arten som eksisterer i dag ... nesten var alle moderne phyla raskt kommet til stede, fullt utstyrt med øyne så langt som man kan fortelle fra fossilene" og under den kambriske eksplosjonen "virker noe bemerkelsesverdig å ha skjedd ... en rik fauna av makroskopiske dyr utviklet seg, og mange av dem hadde store øyne. ' (12) Sir Steward Duke-Elder, den fremtredende oftalmologen ved sin død i 1979, anerkjente det plutselige utseendet til det perfeksjonerte virveldyrøyet, og noterte:
Den pussige tingen om utviklingen av vertebratøyet er imidlertid den tilsynelatende umiddelbarhet av utseendet og utbygging av dets strukturer i sine tidligste kjente faser. Det er ingen lang evolusjonær historie som vi har sett blant virvelløse dyrs øyne, hvor en intracellulær organell passerer inn i en et cellulært og deretter et multi-cellulært øye, som oppnås ved prøving og feiling langs ulike ruter en stadig økende grad av kompleksitet. Innenfor en rekke av virveldyr viser øyet ingen fremgang med økende differensiering og perfeksjon som det ses i hjernen, øret, hjertet og de fleste andre organer. I essensen er fiskens øye så komplisert og fullt utviklet som en fugls eller manns.' (13)
Biokjemiske studier har vist at menneskelinsen inneholder
'... proteiner som ligner de som finnes i syklostomene (hagfishes (kjeveløs fisk) og niøye) som er de levende etterkommerne til Agnatha, som stammer fra virveldyr for 450 millioner år siden. Dermed har disse studiene bekreftet oppfatningen om at vertebratøyet, og spesielt linsen, har endret seg svært lite i løpet av evolusjonen. ' (14)
Bevis for øyets utvikling fra levende dyr
Bare omtrent en tredjedel av alle animalske phyla inneholder arter med skikkelige øyne, en tredjedel inneholder arter med lysfølsomme organer, og en tredjedel har ingen måte å oppdage lys, men mange kan oppdage varme. (15) Ikke desto mindre, av de dyrene med øyne, både vertebrater og de fleste vertebrater, eksisterer et enormt utvalg av øyeformasjoner, plasseringer og størrelser. (10) Øyeeplse diameteren varierer fra mindre enn en tiendedels millimeter i visse vannlopper til 370 mm i gigantisk blekksprut. (16) Øyeplassering varierer også, alt fra den vanlige binokular-visjonen som brukes av de fleste pattedyr til det bevegelige øye på hver side av hodet som brukes av mange øgler/firfisler.
Antall øyne i ett dyr kan også variere fra 0 til åtte. I edderkopper alene, varierer tallet fra null til åtte, alltid eksisterende i to par. Noen øyne inneholder både en linse og en netthinne-liknende struktur i en enkelt celle. (17) Et komplekst teleobjektiv-øye ble identifisert i kameleon i 1995. Grunnen til at så mange mønstre eksisterer er at øynene må tjene meget forskjellige livsformer som lever i svært forskjellige miljøer. Dyr lever i jorda, innsiden av andre dyr, i luften, på land, i saltvann og i ferskvann. Videre varierer dyrene i størrelse fra en vannloppe til en hval.
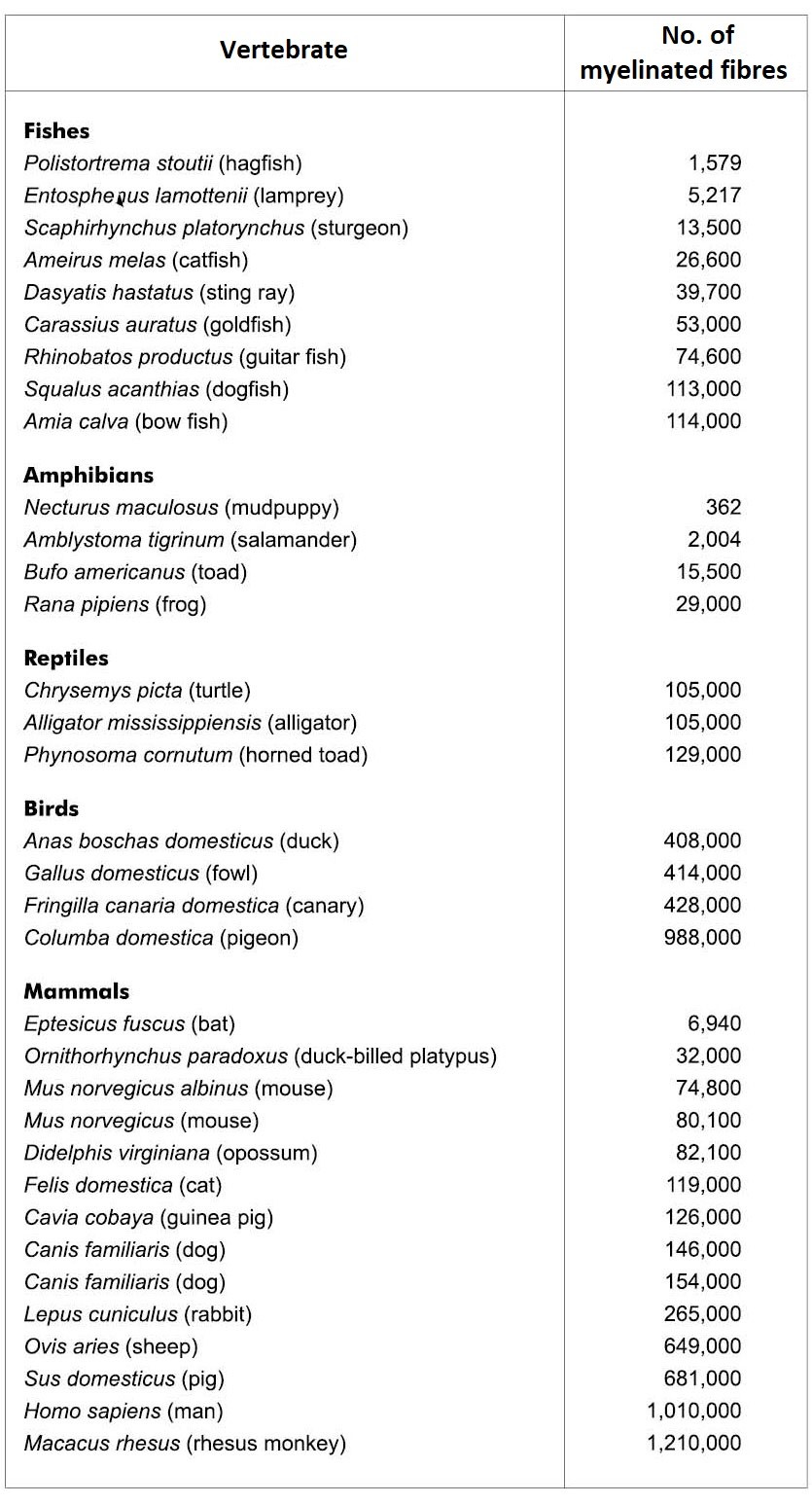
Gjennomsnittlig antall myelinerte fibre i optisk nerve hos utvalgte vertebrater.
Tabell 1. Gjennomsnittlig antall myelin fibre i optisk nerve hos utvalgte vertebrater. Legg merke til den enorme forskjellen i hver kategori. For eksempel varierer fugler fra 408 tusen til 988 tusen, pattedyr fra 7000 til 1,21 millioner. (Fra Cousins (50) ).
Selv om mange forskjellige, meget forskjellige, øyne er kjent, eksisterer det ikke direkte bevis for å understøtte øyets utvikling og tilhørende strukturer. Videre er mye bevis i strid med slike evolusjonære antagelser. Legg merke til i tabell 1 at antall myelinerte fibre i optisk nerve ikke korrelerer med antatt evolusjonær utvikling. En due har nesten like mange fibre som et menneske. Mange fugler, som ørn og hauk, har utmerket syn, men har halvparten så mange fibre som en vanlig gris.
Et annet eksempel er visuelle pigmenter. Den antagelig høyeste, mest utviklede livsformen, de høyere primatene, har bare to kjegle-fotoreceptorer, blå og grønn, men fugler har totalt seks pigmenter: fire kjegle-pigmenter pluss pinopsin (et pineal photoreseptivt molekyl) og rhodopsin for svart og hvitt syn. (12,18) Sagt annerledes har kyllinger, mennesker og mus alle rhodopsin-pigmentet; mus har i tillegg blå og grønn; mennesker har blå, grønn og rød; og fugler har disse tre pigmentene pluss fiolett og pinopsin. For hver farge som mennesker oppfatter, kan fugler se veldig forskjellige flere farger, inkludert ultrafiolett lys. Fugler bruker infrarødt lys (som vi føler som varme) for nattesyn, slik at de raskt kan se sine unger i et tett, mørkt tre.
Muligheten for å klassifisere øyne i levende dyr fra enkle til komplekse enkle typer som finnes i enkle dyr og komplekse typer i komplekse dyr (som vi vil vise ikke kan gjøres), gir ikke bevis for et evolusjonært forhold. Et primært problem er at dette forsøket bare er basert på øye-egenskaper slik de for tiden eksisterer. Historisk øyeutvikling kan ikke bevises ved å notere en rekke eksisterende øyne fra enkle til komplekse og deretter argumentere for at kompleksitet utviklet seg fra det enkle fordi evolusjonen krever at alle eksisterende øyne har en like lang evolusjonær historie.
Darwinister må bestemme øye design fra hvilke eksisterende øyne faktisk har nedkommet, den ene fra den andre over tid. Hertug-Elder og Darwin (1872) foran ham var ikke i stand til å gjøre dette, men de tilbød sin liste over øyne av varierende kompleksitet som evidens for evolusjon. Cousins skrev:
'... Den viktige betydningen av dette kravet til evolusjonsteorien ble fullt forstått av Darwin, som uttalt at i å lete etter graderingen som et organ i noen art har blitt perfeksjonert, burde vi se på deres lineære forfedre. Faktisk burde vi det; selv om han selv ikke kunne gjøre det. Det er villedende for leseren å skape en serie som begynner med øyeflekker som sett i encellede organismer og kalle dem, slik Duke-Elder gjør (1958), det tidligste utviklingsstadiet.' (19)
Croft konkluderte med at kravet om at vi kan stille opp øyne i en evolusjonær rekkefølge fra veldig enkelt til svært komplekst, er falskt fordi forskning på øyets utviklingshistorie i vidt forskjellige arter finner:
'... dem bemerkelsesverdig like. Faktisk er de grunnleggende egenskapene til øyet i forskjellige virveldyr veldig mye de samme til tross for store variasjoner i deres livsstil og tilpasning til habitat. Videre, i motsetning til andre organer som hjertet, er det ingen lang evolusjonær historie med øyet. I hovedsak er øyet av en vannsalamander så komplekst og fullt utviklet som en manns. (11)
Sinclair konkluderte også med at vertebrater og de fleste hvirvelløse dyr, inkludert insekter og klassen av bløtdyr (inkludert blekksprut og tiarmet blekksprut), alle har øyne med vanlige visuelle elementer, inkludert 'et lignende fotoreceptor-design', men likevel har en merkbar 'forskjell i utseende'. (10)
Kilden til øyets design og evolusjon, postulerer darwinister, var en rekke fordelaktige mutasjoner som måtte skje i passende samling for å kunne produsere settet med strukturer som kreves for at øynene skal fungere. Det nye mutasjonssettet, argumenterte Darwinister, resulterte i en overlegen struktur i forhold til den gamle, og dette nye og bedre øye forbedret dyrenes evne til å konkurrere mot andre livsformer. Noen av de mange problemene med denne konklusjonen ble notert av Grassé i sin diskusjon om Myrmelion (insektfamilie) anatomi:
"Har du noen gang sett en mutasjon samtidig påvirke to separate komponenter i kroppen og produsere strukturer som passer perfekt til hverandre? ... har du noen gang sett tre, fire eller fem samtidige mutasjoner med matchende strukturer som produserer koordinerende effekter? ... Dette er viktige spørsmål som krever et svar. Det er ingen måte å komme seg rundt, eller unngå problemet. Hver biolog som vil vite sannheten, må svare på dem, eller bli betraktet som sekteristisk og ikke en forsker. I vitenskap er det ingen "årsak" som trenger bli forsvart, bare bare sannhet som skal oppdages. Hvor mange tilfeldigheter vil det ta å bygge denne ekstraordinære skapningen [ Myrmelion formicarius ] '? (20)
Et organ som ikke hjalp dyrets overlevelse ville bruke knapp energi, næringsstoffer og kroppsrom, og hvis organet ikke ble brukt, ville det være høy risiko for problemer som infeksjon. En øye modifikasjon ville ikke bli valgt før det ikke bare var funksjonelt, men produserte et system som demonstreres bedre enn det eksisterende organet. Først da kunne naturlig utvalg fungere til å velge mellom eksisterende variasjoner for å perfeksjonere organet utover bare funksjonell effektivitet.
Tabell over sammenheng
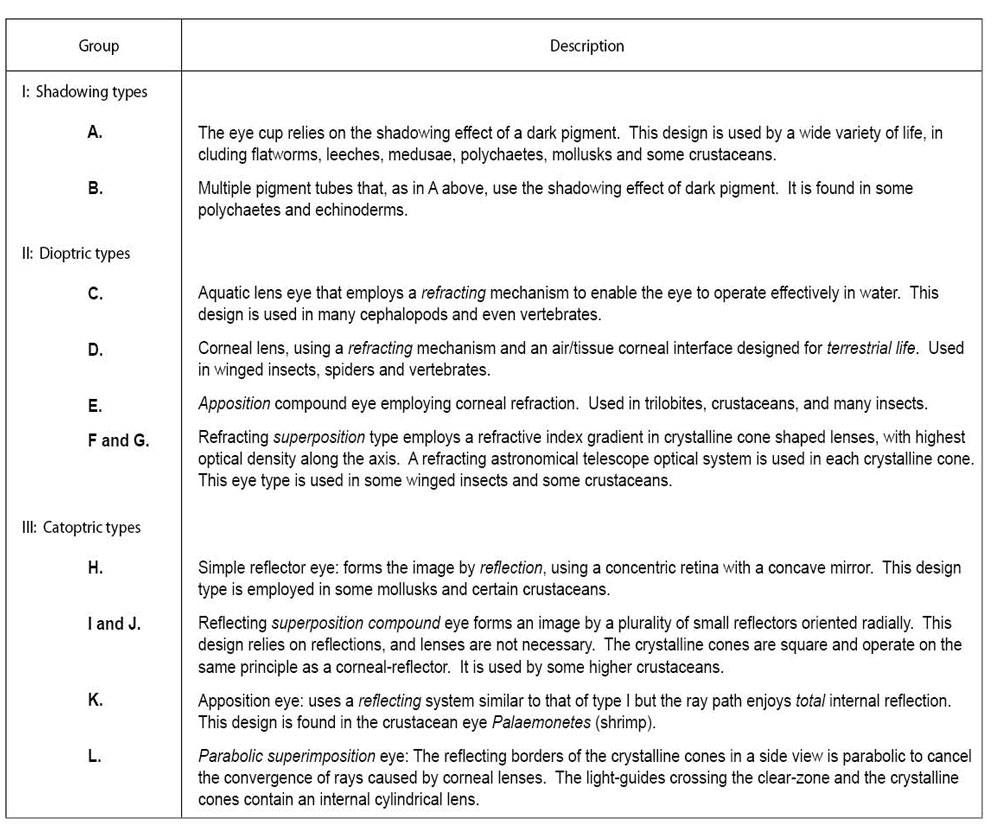
Land og Nilssons mye brukte klassifikasjonssystem for øyeutforminger.
Tabell 2. Land og Nilsson er mye brukt klassifikasjonssystem for øye design. Andre systemer brukes også i dag, noe som illustrerer problemene ved å arrangere øyeutforminger i hierarkier. Vær også oppmerksom på at Land og Nilsson-systemet ikke viser et klart, enkelt og komplekst designhierarki. (Fra Land og Nilson 12 ). Klikk for større visning.
Avansert øye design
Det finnes mange slags øyne, og det er mange ordninger for å klassifisere dem. Det mest grunnleggende klassifiseringssystemet grupperer alle øynene i fire klasser. Den første er kameratypen eller "enkelt" øye, slik som det finnes hos mennesker, som bruker et fokuseringssystem for å projisere et enkelt, skarpt bilde på netthinnen. Den andre typen er fastfokusforbindelsestypen (figur 1) som bruker flere separate brytningsenheter kalt ommatidia, slik som brukt av trilobitter og fluer. Den tredje typen er et skanneøye som bygger et bilde som et fjernsynskamera, som det brukes i den lille marine krepsdyr copiliaen, som hos huner tar opp mer enn halvparten av kroppen. (21) Den fjerde typen er det komplekse øyet, som finnes i cephalopods og visse avanserte vertebrater, bestående av hornhinnen, iris, linser, retina og mange tilbehørskonstruksjoner. (22)
Denne divisjonen skjuler mange store forskjeller: Noen reker har et kombinert enkelt og sammensatt øye, som faktisk er en tredje grunnleggende øyetype, ikke en overgangsform. Dette divisjonssystemet overforenkler også variasjonen som eksisterer fordi "minst elleve forskjellige optiske metoder for å produsere bilder" nå er kjent. 23 Klassifikasjonssystemet som ble benyttet i dette papiret ble utviklet av Land and Nilsson (2005) og er gitt i tabell 2 (se også figur 2).
Problemer med klassifisering
Land og Nilssons mye brukt klassifikasjonssystem for øyeutforminger.
Figur 2. Illustrasjon av Land og Nilssons klassifikasjonssystem for øyemønster. Øyedesign A-L er beskrevet i tabell 2. (Fra Land og Nilson (12)).
Bilde 3. Lysbrytning i ulike øyne
 Problemer med øye-klassifikasjon
Problemer med øye-klassifikasjon
Legg merke til at den mest logiske klassifiseringen av øyetyper er til en slags evolusjonær klassifisering fra enkle til mer komplekse, men denne listen gir seg ikke særlig godt til et hierarki som postulert av Darwin. Faktisk er det umulig å ordne bare de 10 grunnleggende øyeformene som brukes i Land og Nilsson-systemet fra enkelt til komplisert.
For eksempel ser typene A, B, C, D, E, F, H og I ut som i kompleksitet, og typene G og J virker mer komplekse, men finnes i lavere livsformer (i enkelte bevingede insekter og krepsdyr). I Landes klassifikasjon finnes den "enkleste" typen (A) og den mest komplekse typen (J) både i krepsdyr (krepsdyr design i gruppe A, E, F, G og H og bløtdyr i gruppe A og H) . Nesten identiske optiske konstruksjoner finnes i svært 'tydelig ikke-relaterte dyr' som fisk og blekksprutter. (24) Land--listen grupperer bare de grunnleggende øyeformene og optiske systemene, ignorerer utformingen av retina-celler, de mange støttende celler, (som ganglionceller, amakrine celler, horisontale celler og bipolare celler), de andre nervesystemets komponenter, inkludert optisk nerve -, og det optiske system-behandlingssenteret, slik som optiske hjerne-lappen.
Å bruke disse kriteriene ville medføre enda flere problemer i forsøk på å produsere et hierarki, fordi prosess-systemet alltid er mye mer komplekst enn lysoppsamlings-systemet, og plasserer alle kjente øye-systemer på det øvre nivået av Darwins system. Selvfølgelig var Darwin ikke klar over visjonssystemets enorme kompleksitet eller variasjon, og han var heller ikke klar over kompleksiteten til de mange tilbehørssystemene og prosess-strukturer som hjernen.
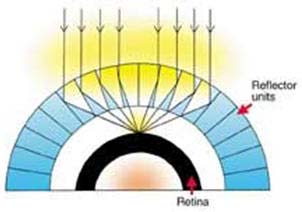
Problemene med å produsere et enkelt til komplekst hierarki er illustrert av det faktum at de ti typene også ofte er arrangert i fire grunnleggende øyeutforminger: det holokroale øyet, overlegningsøyet, det schizokroale øyet og det menneskelige apposjons-forbindelsesøyet . Alle disse grunnleggende øyeformene krever et system med fokuseringsoppløsning og et komplekst nevrologisk behandlingssystem som gjør det mulig for betrakteren å gi mening om den store massen av stadig skiftende signaler sendt av netthinnen eller andre lysfølsomme celler via den optiske nerven til hjernen effektivt og raskt.
Bilde 4. Godt fargesyn?
 "Til tross for flere tiår med forskning har vi fremdeles bare begrenset forståelse av hvordan synet egentlig virker", noe som gjør det vanskelig å produsere både konsistente klassifikasjonsordninger og hierarkier i et forsøk på å postulere en fornuftig evolusjonær fylogeni. (23) Vi har en ganske god forståelse av selve øyestrukturen, noe som tillot konstruksjon av klassifiseringen ovenfor. I motsetning til evolusjonære forventninger kan øynene til fylogenetisk fjerne livsformer være svært 'liknende i et stort antall detaljer.' (16)
"Til tross for flere tiår med forskning har vi fremdeles bare begrenset forståelse av hvordan synet egentlig virker", noe som gjør det vanskelig å produsere både konsistente klassifikasjonsordninger og hierarkier i et forsøk på å postulere en fornuftig evolusjonær fylogeni. (23) Vi har en ganske god forståelse av selve øyestrukturen, noe som tillot konstruksjon av klassifiseringen ovenfor. I motsetning til evolusjonære forventninger kan øynene til fylogenetisk fjerne livsformer være svært 'liknende i et stort antall detaljer.' (16)
Ironisk nok eksisterer det største utvalget av øye-utforming, ikke bare i struktur, men også i antall og plassering, ikke blant virveldyr som Darwinismen forventer, men blant de såkalte "primitive" hvirvelløse dyr. (16) Virveldyr har også øyne som i noen henseender er bedre enn hos virveldyr. Et eksempel er de halvkuleformede øynene til de fleste fluer og andre insekter som, i motsetning til menneskelige og fleste virveldyr-øyne, produserer et bilde som i stor grad er fri for sfærisk forvrengning. (25) Menneskelige øyne har betydelig perifer bildeforvrengning, men sfæriske øyne danner et skarpt bilde i alle retninger. Imidlertid har mennesker ikke skarpt perifert syn fordi dette er funksjonen til den sentrale retinaen som kalles makulaen. Vårt perifere syn er for gjenkjenning av lys og bevegelse som utløser fikseringsrefleksen for å snu øynene mot stimulansen.
Et annet problem i teorien om at øye-utformingene representerer en evolusjonær sekvens er at øynene fra de tre store phyla (virveldyr, leddyr og bløtdyr) stammer fra forskjellige vev og er radikalt forskjellige. (26) Derfor konkluderte evolusjonister at de har separate evolusjonære historier, og de mange likhetene som eksisterer skyldes antatt evolusjonær konvergens. (26) I utgangspunktet, "vi vet ikke hvordan det kunne ha utviklet seg, så det må ha utviklet seg om og om igjen." Øyeforskjellene vil skyldes ulike behov og forhold for hver organisme og dens habitat, uavhengig av evolusjonær sammenheng. Enda et annet problem er at bevis for øyeutvikling styrker konklusjonen at de fleste av disse øyeformene må ha utviklet seg "i en kort periode under Kambrium." (17)
Det enkleste øye
Darwinister hevder ofte at primatøyet er den mest utviklede, men mange feilbenevnte 'primitive' øyne har fordeler over oss. For eksempel kan det menneskelige øyet registrere opp til 60 bilder per sekund; en lite bi omtrent 300 per sekund. Av denne grunn kan bier se langt bedre mens de beveger seg raskt. Filmbilde-standarden (24 bilder per sekund), til en bi, vil bli sett på som en serie stillbilder. For mennesker er rammene uskarpe, noe som gir en illusjon av bevegelse. Denne design-innovasjonen i såkalte primitive dyr er mer komplisert enn den tilsvarende strukturen i det menneskelige øye.
Den enkleste øyetypen er kjent som ocellus, et multi-cellulært øye bestående av fotoreceptor-celler, pigmentceller og nerveceller for å behandle informasjonen - er trinn 4 i Darwins liste. (27) Det mest primitive øyet som oppfyller definisjonen av et øye, er den lille - om størrelsen på hodet til en pin-mikroskopisk marine krepsdyrkopipod copilia . Bare hunnene har hva Wolken og Florida kaller "bemerkelsesverdige øyne som utgjør mer enn halvparten av sin gjennomsiktige kropp." (28) Hevdet å være en kobling mellom en øyeflate og et mer komplekst øye, har den to utvendige linser som rasterer som et skannings-elektronmikroskop for å samle lys som behandles og deretter sendes til hjernen.(29) Den har retinale celler og et øye som er analogt med en superposisjon-type (ommatidium) av sammensatte øyne. (30) Dette, det mest primitive sanne øyet, er kjent på stadium 6 i Darwins evolusjonære hierarki!
Bilde 5. Lysbrytning i hummer øyne
 Visuelle celleforskjeller
Visuelle celleforskjeller
Evolusjonen vil forutsi at det mer avanserte øyet, jo mer detaljert det kan hente, en faktor relatert til antall visuelle celler. Dette er ikke det som ofte blir funnet. I et "enkelt" visuelt system (hjerne og retinaer) er det minste antall visuelle celler funnet i pletodontidsalamander, T. narisovalis , som bruker ca. 65.000 celler for hele det visuelle hjernesenteret og 60.000 for netthinnen alene. Dette "ekstremt lave" antall celler brukes ikke fordi dyret er primitivt, men fordi det har et veldig lite hode, øye og hjerne pluss relativt store celler. (31) De legger til at den minste, utbredte salamander, T. pennatulus (som er mye mindre enn T. narisovalis ), har om lag 94 000 visuelle celler og omtrent det samme antall retinale celler. Til sammenligning inneholder hjernens visuelle sentre av frosken S. limbatus omtrent 400.000 celler. Dette illustrerer det faktum at evolusjon ikke kan argumenteres
'... ved å hevde at øyet kan bygges opp gradvis fra en enkelt lett hudfølsom hud gjennom ulike stadier, og langsomt nå kompleksiteten til virveldyr-kameraøyet. ...
Saken for utviklingen av virveldyrøyne eller til og med en lysfølsom hudplast ... må gjøres med hensyn til hele organismens kompleksitet, i det minste i den grad kompleksiteten støtter visjon (selv i minst komplisert form). Av denne grunn bør debatten ikke handle om øyets utvikling, men om synets utvikling, og syn er alltid synet til en bestemt type levende dyr, en levende helhet hvor den integrerte aktiviteten og erfaringen med å se, selv i sin enkleste form, kan finne sted. ' (32)
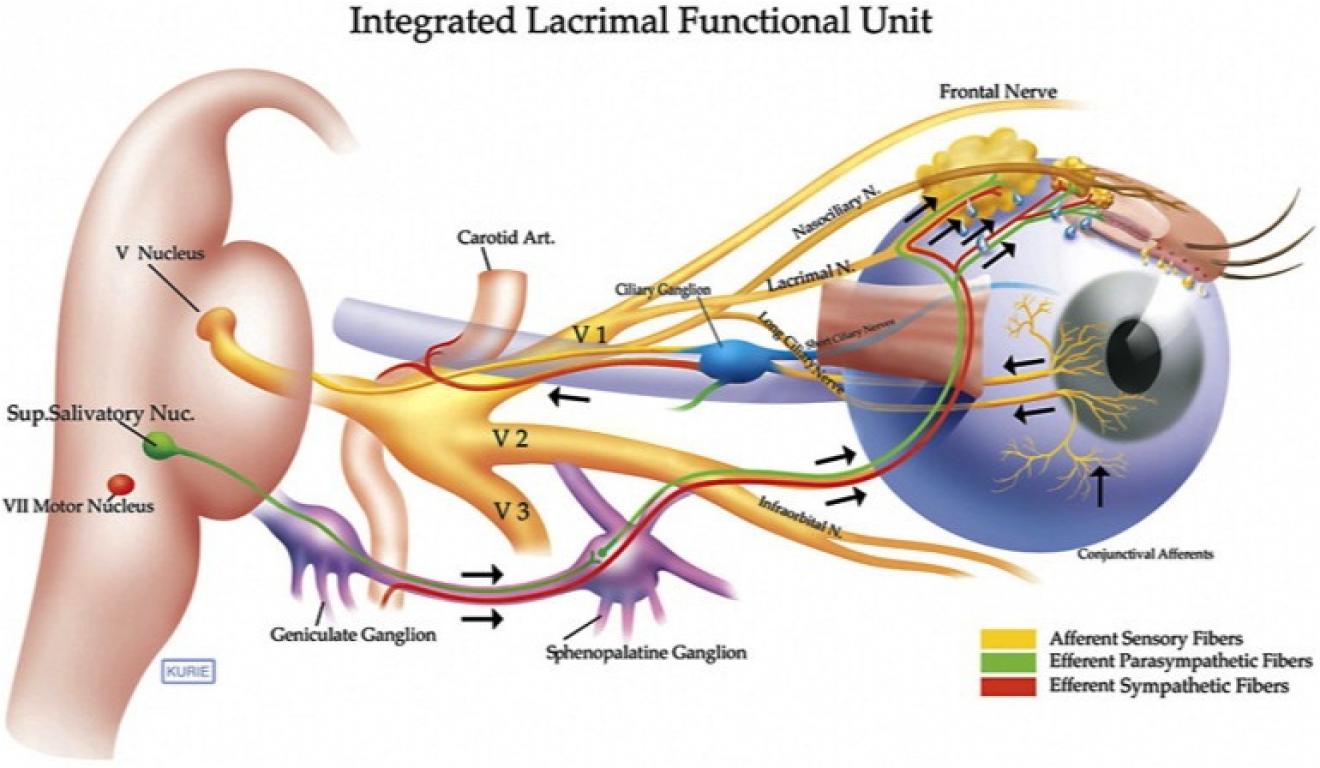
I tillegg til antall celledifferanser eksisterer også foto-reseptorcelle forskjeller. Cellene som gir membranoverflaten for opsin-molekyler, kan enten være ciliary eller microvillar strukturer. Microvilli-typen dominerer hos virvelløse dyr og cilia-typer hos virveldyr. Selv fysiologiske responser varierer mye. Lys forårsaker mikrovilløse reseptorer av leddyr og bløtdyr å depolarisere, men forårsaker at de ciliære reseptorene hos virveldyr hyperpolariserer. Virvelløse dyr bruker inositoltrifosfat til fotodetransduksjon i det andre messenger-systemet, mens virveldyr-fotoreseptorer bruker syklisk guanosin 5'-monofosfat (GMP). Selv om opsin er nøkkelmolekylet som brukes til å oppdage lys i både vertebrater og hvirvelløse dyr, er regenererings-mekanismer (reisomerisering) av kromofor/opsin-systemet 'dramatisk forskjellig blant dyrerekkene'. (33) Andre viktige forskjeller er virvelløse dyrs øyne som dannes fra hudens overflate av ektoderm og virveldyrs øyne som er dannet fra den neurale ektodermen. (34)
Et annet problem for evolusjon er at minst 11 forskjellige optiske metoder brukes til å produsere bilder. For en type å utvikle seg til en mer avansert type krever mellomliggende trinn som er mye verre eller ubrukelige i forhold til eksisterende design. Dette ville gjøre en endring i det hovedsak dødelig for dyr som er avhengige av synet. ' (35) For eksempel har de avanserte stavene og kjeglene i "primitive" dyr og mangelen på bevis for deres utvikling motivert noen til å konkludere med at det "grunnleggende tetrachomatiske systemet utviklet seg veldig tidlig i utviklingen av vertebrater". (36) Videre eksisterer det ingen fremgang fra enkle til komplekse fotoreseptorer, men bare "fire spektralt adskilte klasser av kjeglepigment kodet av forskjellige opsin-gener" finnes i den naturlige verden. (37)
Bilde 5. Norske navn --menneskeøyets oppbygning
 Evaluering av gener involvert i øyeutvikling
Evaluering av gener involvert i øyeutvikling
Omvendt, likheter, som det faktum at noen av genene som er involvert i øyeutvikling, er svært likt i de fleste dyr, argumenterer for en enkelt evolusjon av øyet. Likevel er vanskeligheten med øyeutvikling så stor at øynene som hypotese av enkelte forskere har utviklet seg selvstendig minst 40 og så mange som 65 ganger. (38) Som Fernald noterer, for tiden, "vi vet ikke om øynene oppstod en eller flere ganger, og faktisk er mange funksjoner i øyeutviklingen fortsatt forbløffende." (23) En bedre forklaring på det samme genet som brukes av forskjellige dyr (eller planter) er for ressurs-sparing i design av en høyere intelligens.
Virveldyr øyne kunne ikke ha utviklet seg i isolasjon fordi øyedeler ikke har en funksjon som selvstendige enheter. Øyne er en del av svært komplekse, sammenkoblede levende organismer, og øynene er bare en del av visjonssystemet. (39)
En målestokk for å bestemme øyekompleksiteten er antall gener som er involvert i å produsere øyet - jo mer gener som kreves, jo mer komplisert kan øyet være. I den primitive Drosophila har hittil blitt identifisert 501 øye-relaterte gener, eller ca. 3,5% av hele dets genom. (24) Virveldyrs øyne anslås å involvere 7.500 gener bare for å utvikle og regulere retina - eller ca. 30% av hele det menneskelige genom av 25.000 gener. (24)
Visninger av øyeutvikling har flip-flappet
Disse problemene er en del av grunnen til at "syn på øyeutvikling har flip-flappet, til alternativt å favorisere en eller mange opprinnelser." (40) Den markante distinkte ontogenetiske opprinnelsen til øynene i svært forskjellige arter er en grunn til at øynene er postulert for å ha utviklet seg uavhengig 40 eller flere ganger. (40) For eksempel er øynene i mange bløtdyr, inkludert noen blekkspruter som 10-armet og 8-armet blekksprut, bemerkelsesverdig lik virveldyr-øyne. Begge har en hornhinne, en linse, en iris og en netthinne. En av de store forskjellene er, i en, er retina invertert, sammenlignet med den andre. (41)
Evolusjonister forsøker å løse dette problemet ved å anta at den fylogenetiske linjen som førte til bløtdyr splittet veldig tidlig i evolusjonær historie, lenge før øyet hadde utviklet seg. Da postulerer de parallell evolusjon - konkluderer at de to øynene utviklet seg til å være nesten identiske, men var helt uavhengige av hverandre. Av oppmerksomhet er det faktum at det mest 'primitive' kameraøyet kjent (nautilus -'nålehull'-øyet) og det mest avanserte øyet som er kjent, begge finnes i bløtdyr! Bløtdyr som gruppe inneholder en pigment øyeflekk-design, en pigmentkopp (kopulat), en enkel optisk kopp med en 'nålehull'-linse, et øye med en primitiv linse (en murex -marin snegle) og et komplekst øye (blekksprut), sistnevnte som er det "mest forseggjorte" øyet i det virvelløse rike. (42)
Bilde 6. Bildedanningsprosess -signalveier
 Embryonisk opprinnelse av virveldyrs øyne i motsetning til bløtdyr-øyne
Embryonisk opprinnelse av virveldyrs øyne i motsetning til bløtdyr-øyne
En annen stor forskjell finnes i den embryonale opprinnelsen til mange strukturer i virveldyr øyne i motsetning til bløtdyr øyne. F. eks. dannes blekksprut øyne fra en epidermal plate ved påfølgende innfolding, mens virveldyr øyne utvikler seg fra en nevrale plate, og overliggende epiderm danner linsen. Et annet problem for øyeutvikling er at øyet av bare en evolusjonær relatert klasse, virveldyr, 'utvikler seg fra en mangfoldig samling av embryonale kilder gjennom et komplekst sett med induktive hendelser.' (43)
Konklusjoner
Dennett skrev at øyelinsen er "utsøkt godt utformet for å gjøre jobben sin, og ingeniørens begrunnelse for detaljene er umiskjennelig, men ingen designer har formulert det." Han konkluderer med at designet ikke er ekte, men en illusjon fordi evolusjonen forklarer øyet uten behov for en designer. Denne anmeldelsen har vist at evolusjonen ikke forklarer eksistensen av visjonssystemet, men en intelligent designer gjør det. De ledende øye-utviklings forskerne innrømmer at de bare har en viss forståelse for hvordan øynene kan ha utviklet seg. (45) Disse forklaringene skraper ikke engang i overflaten av hvordan et visjonssystem kunne ha oppstått av evolusjon - enn si "når".
Mye uenighet eksisterer om den hypotetiske utviklingen av øynene, og eksperter innser at mange kritiske problemer eksisterer. Blant disse problemene er en forklaring på utviklingen av hver del av visjonssystemet, inkludert linsen, øyeeplet, netthinnen, hele det optiske systemet, synslappen i hjernen og de mange tilbehørs-konstruksjonene. Turner understreket at "det virkelige mirakelet [av visjon] ligger ikke så mye i det optiske øyet, men i prosessen som gir visjon." (46) Alle disse forskjellige systemene må fungere sammen som en integrert enhet for at syn skal oppnås. Som Arendt konkluderer, har øyets utvikling blitt diskutert siden Darwin og diskuteres fortsatt blant darwinistene. (47) For ikke-evolusjonister er det ingen debatt.
Bilde 7. Øyet -sjelens speil?
 Relaterte artikler
Relaterte artikler
Sammenlignings-anatomi av øyet i dyreriket - med innkalt evolusjon -her.
Fiberoptikk i øyet ødelegger ateistisk 'dårlig design' argument -her.
Er vår "inverterte" retina virkelig "dårlig design"? -her.
Øye tilkobling klargjort -her.
Gigant sammensatte øyne, for en halv milliard år siden? -her.
Benekting av 'dårlig design' argumenter (Creation Magazine LIVE! 4-18) -her.
Referanser og notater
1. Sinclair, S., How Animals See: Other Visions of Our World, Henry Holt, New York, p. xi, 1985. Return to text.
2. Darwin, C., The Origin of Species, John Murray, London, p. 186, 1859. Return to text.
3. Darwin, C., The Descent of Man, and Selection in Relation to Sex, John Murray, London, 1872. Return to text.
4. see Shermer, M., The Case Against Intelligent Design, Henry Holt, New York, p. 17, 2006. Return to text.
5. Isaak, M., The Counter-Creationism Handbook, Greenwood Press, Westport, CT, p. 65, 2005. Return to text.
6. Coyne, J., The faith that dare not speak its name: the case against intelligent design, The New Republic, p. 30, 2005. Return to text.
7. Dennett, D.C., The hoax of intelligent design and how it was perpetrated in Brockman, (Ed.), Intelligent Thought: Science Versus the Intelligent Design Movement, Vintage Books, New York, p. 37, 2006. Return to text.
8. Melnick, J., Vision: an evolutionary enigma, Christian Citizen 1(9):26, 1981. Return to text.
9. Moses, K., Fly eyes get the whole picture, Nature 443:638-639, 2006. Return to text.
10. Sinclair, S., ref. 1, p. 9. Return to text.
11. Croft, L.R., The Last Dinosaurs, Elmwood Books, Chorley, Lancashire, p. 57, 1982. Return to text.
12, Land, M.F. and Nilsson, D.-E., Animal Eyes, Oxford University Press, New York, p. 1, 2005. Return to text.
13. Duke-Elder, S.S., System of Ophthalmology. Volume 1: The Eye in Evolution, The C.V. Mosby Company, St. Louis, p. 237-238, 1958. Return to text.
14. Croft, ref. 11, p. 59. Return to text.
15. Land and Nilsson, ref. 12, p. 4. Return to text.
16. Brain, Behavior and Evolution 64(3):141-147, 2004; p. 143. Return to text.
17. Land and Nilsson, ref. 12, p. 15. Return to text.
18. Jager, S., Lewis, J., Zvyaga, T., Szundi, I and Sakmar, T., Chromosome structural changes in rhodopsin from nanoseconds following pigment photolysis, Proceedings of the National Academy of Science 94:8557-8562, 1997. Return to text.
19. Cousins, F.W., The Anatomy of Evolution, Duffett Publications, London, p. 125, 2003. Return to text.
20. Grassé, P.P., Evolution of Living Organisms, Academic Press, New York, NY, p. 163, 1977. Return to text.
21. Wolken, J. and Florida, R.G., The eye structure and optical system of the crustacean capecod, copilia, The Journal of Cell Biology, 40(1):279-285, 1969. Return to text.
22. Arendt, D. and Wittbrodt, J., Reconstructing the eyes of urbilateria, Philosophy Transactions of the Royal Society of London 356:1546, 2001. Return to text.
23. Fernald, R.D., The evolution of eyes, Karger Gazette 64:2-4,
2001; p. 2. Return to text.
24. Fernald, R.D., Casting a genetic light on the evolution of eyes, Science 313:1914-1918, 2006; p. 1914. Return to text.
25. Sinclair, ref. 1, p. xvii. Return to text.
26. Fernald, R.D., The evolution of eyes, Brain, Behavior and Evolution 50(4):253, 1997. Return to text.
27. Arendt and Wittbrodt, ref. 22, p. 1546. Return to text.
28. Wolken and Florida, ref. 21, p. 279. Return to text.
29. Sinclair, ref. 1, p. xiii. Return to text.
30. Wolken and Florida, ref. 21, p. 280. Return to text.
31. Roth, G. and Blanke, J., Vertebrates in McGraw-Hill Yearbook of Science and Technology, McGraw-Hill, New York, 1997. Return to text.
32. Wiker, B. and Witt, J., A Meaningful World: How the Arts and Sciences Reveal the Genius of Nature, InterVarsity Press, Downers Grove, IL, p. 44, 2006. Return to text.
33, Fernald, ref. 16, p. 146. Return to text.
34. Cousins, ref. 19, p. 284. Return to text.
35. Fernald, ref. 23, pp. 2-3. Return to text.
36. Bowmaker, J.K. and Hunt, D.M., Evolution of vertebrate visual pigments, Current Biology 15(13):R484-R489, 2006; p. R489. Return to text.
37. Bowmaker and Hunt, ref. 36, p. R484. Return to text.
38. Fernald, ref. 16, p. 145. Return to text.
39. Breidach, O. and Kutsch, W., The Nervous Systems of Invertebrates: An Evolutionary and Comparative Approach. With a coda written by T.H. Bullock, 1995. Return to text.
40. Fernald, ref. 24, p. 1917. Return to text.
41. McIlwain, J.T., An Introduction to the Biology of Vision, Cambridge, New York, 1996. Return to text.
42. Lovicu, F. and Robinson, M.L., Development of the Ocular Lens, Cambridge University Press, New York, p. 17, 2004. Return to text.
43. Fernald, ref. 23, p. 3. Return to text.
44. Dennett, D.C., Breaking the Spell: Religion as a Natural Phenomenon, Viking, New York, p. 60, 2006. Return to text.
45. Fernald, ref. 16, p. 467, emphasis mine. Return to text.
46. Turner, J.S., The Tinker’s Accomplice: How Design Emerges from Life Itself, Harvard University Press, Cambridge, MA, p. 161, 2007. Return to text.
47. Arendt, D., Evolution of eyes and photoreceptor cell types, International Journal of Developmental Biology 47:563, 2003. Return to text.
48. Mitchell, L., Mutchmor, J. and W. Dolphin, W., Zoology, Benjamin Cummings, Menlo Park, CA, p. 279, 1988. Return to text.
49. Mitchell et al., ref. 48, p. 278. Return to text.
50. Cousins, ref. 19, p. 300. Return to text.
Bibelen erklærer: I begynnelsen skapte Gud himmelen og jorden. 1. Mosebok 1v1
Oversettelse og bilder ved Asbjørn E. Lund
{kind=link}